Capítulo 5.- MUSCULAR
HISTOLOGÍA
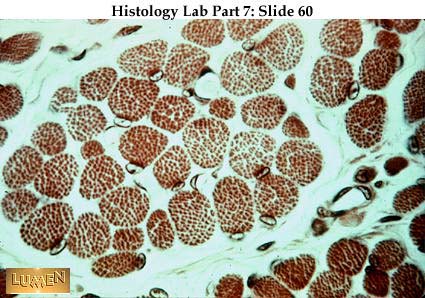
Cada músculo está envuelto por una cubierta de tejido conjuntivo
llamada epimisio. Esta cubierta presenta prolongaciones que penetran
hacia el interior del músculo dando lugar al perimisio y a la división
del músculo en fascículos de diverso tamaño. Finalmente, cada una de las fibras
musculares está envuelta a su vez por una fina lámina de tejido conjuntivo, el
endomisio
{kind=link}
--------------------
Con microscopía electrónica, las miofibrillas aparecen constituídas por
filamentos perfectamente ordenados: unos gruesos, de 110 a 120 Å de diámetro
(miosina) y otros delgados, de unos 60 a 70 Å de diámetro (actina). Los
filamentos de actina se disponen paralelamente y se anclan en unas estructuras
transversales gruesas de 340 a 1300 Å de espesor denominadas bandas Z. Los
filamentos de miosina se disponen centralmente, alternando con los filamentos
de actina. Las zonas más mediales de los filamentos de miosina presentan
uniones de refuerzo con una estructura reticular que en los cortes
longitudinales aparecen como bandas densas o bandas M. La estructura limitada
por dos bandas Z se denomina sarcómero y constituye la unidad funcional
contráctil del músculo estriado
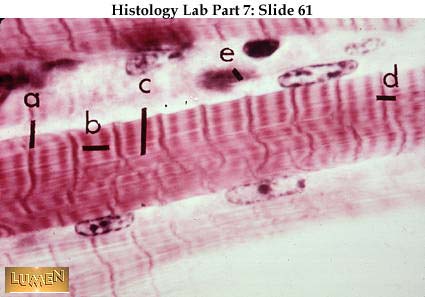
En la diapositiva* enlazada se observa:
a) Línea Z
b) Zona H y banda A
c) Banda M
d) Banda I
{kind=link}

--------------------
Enlaces generales
La actina se dispone en cadenas largas de tipo helicoidal (girando sobre
si misma) denominadas actina F (fibrosa), formadas por monómeros de actina G
(globulosa o elipsoide). Cada siete monómeros de G-actina da media vuelta la
hélice de F-actina

En los surcos de la hélice que conforma la actina, hay moléculas de una
proteína fibrosa y alargada denominada tropomiosina.
Cerca del extremo de cada molécula de tropomiosina existe un complejo
proteico denominado troponina.
Esta troponina está formada por tres subunidades, la mayor (Tn-T) tiene
capacidad para unirse a la tropomiosina; la Tn-I es capaz de inhibir la
actividad ATP-asa de la actomiosina; y la Tn-C, la más pequeña, es capaz de
ligar cuatro iones de calcio
La miosina es la unidad fundamental de los filamentos gruesos. Alrededor
de dos tercios de las proteínas de los músculos esqueléticos son miosina. Cada
filamento de miosina está formado por unas 200 moléculas de miosina alineadas.
La molécula de miosina es un hexámero compuesto por dos cadenas polipeptídicas
con un alto peso molecular (cadenas pesadas), y otras cuatro cadenas de bajo
peso molecular (cadenas ligeras). Cada molécula de miosina, por tanto, se
compone de dos partes, meromiosina ligera y pesada, y cada filamento de miosina
está formado, como hemos dicho, por unas 200 moléculas de miosina
La cabeza de la miosina, denominada región S1 es la responsable de la actividad enzimática y
química, lo que conduce a la contracción muscular. Esta región tiene un sitio
de unión al ATP, lo que es imprescindible para la energía necesaria en la
contracción. La región S2 sirve
como unión flexible entre las regiones de la cabeza y la cola. En esta zona se
produce la flexión de la cabeza de la miosina que da lugar al deslizamiento de
la miosina sobre la actina y al acortamiento muscular que se produce en la
contracción.
En la región S1 existen dos cadenas peptídicas,
una de ellas se denomina esencial porque es necesaria para el funcionamiento de
la miosina, la otra, que se denomina reguladora, puede fosforilarse durante la
actividad muscular y participa en la regulación de la contracción muscular.

FASE ELÉCTRICA
Enlaces generales
Entre el exterior y el interior de la fibra muscular hay una diferencia
de potencial de –90 mV (debido a la diferencia de concentración iónica entre
ambas partes de la membrana celular).
Cuando llega un potencial de acción a la placa motora se libera
acetil-colina, neurotransmisor que se une a receptores de tipo nicotínico que
existen en la célula muscular. Al unirse la acetil-colina a su receptor se
produce un cambio en su posición, lo que genera un poro por el que pasan iones
modificando la situación de equilibrio Na+-K+ . El
potencial de membrana pasa a ser de hasta + 40 mV. Esta variación del potencial
de membrana origina circuitos locales entre la zona activa y la inactiva,
produciendo corrientes eléctricas registrables mediante electrodos
extracelulares (fenómeno base de la electromiografía). La despolarización de la
membrana se propaga, entonces, con una velocidad de 4-5 m/sg hacia los dos
extremos de la fibra muscular.

---------------------
FASE MECÁNICA
a) Los filamentos de actina y miosina se encuentran disociados
b) La presencia de Ca++ inducida por su salida del retículo
sarcoplásmico, produce la unión de la actina a la miosina, ya que el calcio se
une a la troponina y permite que la actina se fije a la cabeza de la miosina.
El ATP que está disociado y formando un complejo con la miosina (M*ADP+Pi),
interacciona con la actina, rota la cabeza de miosina y se libera el ADP y el
Pi.

c) La rotación de las cabezas de miosina, unidas temporalmente a los
filamentos finos, tira de éstos, produciendo un movimiento que se denomina golpe de potencia, acortando el
sarcómero.
d) Sigue siendo rígida la unión entre actina y miosina, y se precisa ATP
para unirse a la cabeza de la miosina y formar el complejo M*ADP+Pi. El calcio
vuelve al interior del retículo sarcoplásmico.


Enlace imagen de la contracción
muscular
{kind=link}
ACOPLAMIENTO ELECTRO-MECÁNICO
Para producir una fuerza mecánica eficaz, las distintas partes de la
fibra muscular deben desarrollar su tensión simultáneamente. De hecho, si el
potencial de acción muscular utilizara una sustancia que difundiera desde la
membrana hacia el centro de la fibra, el retardo en la puesta en marcha sería
demasiado lento y provocaría contracciones separadas e inútiles. Para llevar la
despolarización inicial producida en la membrana, al interior, se pone en
marcha un sistema de túbulos T que permite la salida del Ca++ almacenado en su interior al sarcoplasma.
La concentración del calcio en el citoplasma de la fibra muscular será
el factor determinante del estado de contracción o de relajación en que se
encuentre el músculo. Cuando existan en el sarcoplasma concentraciones
superiores a 10-5 M se producirá contracción.
Cuando cesa el estímulo nervioso, descienden los niveles de Ca++ en el citoplasma debido a la acción de una
ATPasa calcio-dependiente situada en la membrana del retículo sarcoplásmico,
que bombea continuamente calcio del sarcoplasma al interior de las vesículas
del retículo sarcoplásmico, hasta conseguir que las concentraciones de Ca++ desciendan por debajo de la cifra citada.

----------------------
UNIDAD MOTORA
La unidad motora está constituida por la motoneurona y las fibras musculares
que inerva.
Las características generales a destacar de la unidad motora son:
a) Todas las fibras de una unidad motora tienen homogeneidad en lo
referente a la contractibilidad y a las propiedades metabólicas.
b) Las fibras de una unidad motora están distribuidas en una amplia área
de la sección transversal de músculo.
c) Las fibras que pertenecen a una misma unidad motora raramente están
situadas inmediatamente próximas las unas de las otras.
d) La localización central de la unidad motora viene determinada por la
mayor densidad de fibras de dicha unidad, declinando la condensación de las
mismas conforme nos alejamos de dicha localización central.
En el hombre, parece que sólo son de 15 a 30 las unidades motoras que
están contenidas en 5-10 mm2 de área
de sección transversal.
El número de fibras por unidad motora va a ser determinado por la
precisión, exactitud y coordinación que va a requerir el movimiento. Ejemplo:
el número de fibras por unidad motora es de 110 para los músculos de los dedos,
y de 1.720 para los gemelos (es decir, para más precisión mayor cantidad de
unidades motoras, a las que corresponde menor número de fibras por unidad).
Cuando un impulso nervioso alcanza la placa motora, se libera acetil
colina, despolarizándose la membrana de la fibra. Se producirá un potencial de
acción que se propaga por la fibra a una velocidad de 5 m/sg. De aquí se
desprende que la totalidad de una fibra y todas las fibras de una unidad motora
se ponen en acción casi instantáneamente.
Ley del todo o nada: Un único estímulo de intensidad suficiente da
origen a una contracción de la fibra muscular. Después de un corto período de
tiempo, la tensión desarrollada disminuye, y la fibra se relaja.
Efecto de sumación: Si el nervio motor es estimulado repetidamente, y el
segundo estímulo alcanza la fibra antes de su total relajación, se contrae de
nuevo, pero como esta contracción comenzó a un nivel de tensión más alto, la
tensión del segundo estímulo será considerablemente mayor que la del primer
estímulo.
Tetanización: En altos niveles de estimulación, el músculo no se relaja
antes de la siguiente contracción, y las fibras musculares entran en tetania,
es decir, hay una fusión mecánica de las contracciones. A mayor nivel de
estimulación habrá una mayor tensión desarrollada por la fibra*
{kind=link}
FIBRAS TIPO I (lentas u oxidativas). Poseen un tipo de miosina compuesta
por cadenas pesadas con actividad enzimática ATPasa de baja velocidad máxima,
la isoforma MHC I (Myosin Heavy Chain I). Son las fibras predominantes en los
músculos antigravitatorios y los usados en la locomoción (sóleo). Asimismo, las
cadenas ligeras esenciales de la miosina tienen en estas fibras características
propias (son las isoformas MLCs (Myosin Light Chains))
Las demás proteínas
como la tropomiosina tienen también características propias.
En cuanto al complejo
de la troponina, tanto la C, como la T y la I presentan isoformas
características de este tipo de fibra.
La actina no presenta
isoformas distintas según el tipo de fibra musculoesquelética.
Finalmente, la línea Z
es más ancha en este tipo de fibras y el sistema contráctil se dispone en
miofibrillas que son mas escasas que en las fibras tipo II, con lo que queda
una mayor proporción de sarcoplasma libre.
Por otro lado, las
fibras tipo I presentan un retículo sarcoplásmico menos abundante que las tipo
II, los túbulos T presentan menor superficie de membrana y suponen un menor
porcentaje del volumen celular que en las tipo II.
Referente al
metabolismo energético, las fibras tipo I obtienen su energía,
fundamentalmente, del ciclo de ácidos tricarboxílicos y de la fosforilación
oxidativa. Tienen, por tanto, mayor número de mitocondrias y mayor reserva de
triglicéridos en pequeños depósitos intramusculares.
FIBRAS TIPO II. Poseen un tipo de miosina compuesta por cadenas pesadas
con actividad enzimática ATP-asa que varían entre las isoformas MHC IIa y MHC
IIx, lo que subdivide a este tipo de fibras en IIa y IIx, siendo la de mayor velocidad la IIx.
También, el complejo de
la troponina, tanto la C, como la T y la I presentan isoformas características
de este tipo de fibra.
Los túbulos T
representan un mayor volumen respecto al volumen celular total que en tipo
anterior de fibras. El retículo sarcoplásmico está mucho mas desarrollado.
Durante el ejercicio físico se reclutan las fibras tipo I en primer
lugar, continuando las IIa seguidas de las IIx. Si la contracción es muy
intensa se activan todos los tipos de
fibras.
Las propiedades de la miosina que
dan la diferencia entre las contracciones rápidas y lentas de las unidades
motoras se desarrollan durante la etapa fetal tardia o en un periodo postnatal
temprano
Existen todo tipo de formas de transición
entre unas fibras y otras en función de las distintas concentraciones de las
isoformas.
La actividad neuromuscular se gradúa sobre la base de un orden fijo de
movilización de la reserva disponible de unidades motoras. Cuanta mas fuerza se
necesita para ejecutar un cierto movimiento, más unidades motoras se movilizan.
Las primeras fibras que se movilizan son las ST, si la resistencia aumenta, se
reclutan fibras FTa.

---------------------
TIPOS DE CONTRACCIONES MUSCULARES
Según la longitud del músculo:
- Concéntrica.- Se produce acortamiento y, por tanto, trabajo en el sentido físico (T = F x S; si no hay desplazamiento, el trabajo en sentido físico es cero). Puede ser isodinámica (isotónica) cuando la fuerza de la contracción se mantiene constante e invariable en todos los ángulos del movimiento, y alodinámica en caso contrario. La mayor parte de las contracciones musculares habituales son alodinámicas.
- Excéntrica.- Cuando simultáneamente con la actividad contráctil, el sistema muscular es estirado. Cuando intentamos mantener la flexión del antebrazo sosteniendo una pesa importante y al ser mayor de lo que podemos soportar nos va venciendo produciéndose la extensión del antebrazo sobre el brazo, estamos realizando una contracción de este tipo. Estas contracciones están mas relacionadas con el dolor muscular (agujetas).
- Isométrica.- Cuando en la contracción no se modifica la longitud del músculo. No existe trabajo en sentido físico.
TIPOS DE
CONTRACCIONES MUSCULARES
Respecto a la velocidad del movimiento:
Isocinética.- Si la
velocidad de movimiento es invariable en el transcurso de la contracción. Son
contracciones logradas con dispositivos especiales, no existen en el movimiento
motor normal en el organismo
Heterocinéticas.-
Cuando si hay variación de la velocidad con la contracción.
En algunas ocasiones
se pueden producir situaciones especiales que dan lugar a un tipo diferente de
contracción. Por ejemplo, si al acortarse el músculo la fuerza que tiene que
ejercer es cada vez mayor, la contracción se denomina auxotónica (tensar la cuerda de un arco, por ejemplo).
----------------------
1.- Las adaptaciones nerviosas acompañan siempre al aumento de fuerza
resultante del entrenamiento contra resistencia, sin embargo la hipertrofia
puede estar o no presente.
2.- Los mecanismos nerviosos que conducen al aumento de fuerza pueden
ser la movilización de mas unidades motoras para actuar sincronizadamente y las
reducciones en la inhibición autogénica de los órganos tendinosos de Golgi.
3.- La hipertrofia muscular temporal es la sensación de hinchazón que se
tiene inmediatamente después de la realización de una sesión de ejercicios. Se
trata de un cierto edema y es de corta duración.
4.- La hipertrofia muscular crónica se produce como consecuencia del
entrenamiento repetido contra resistencia y es el reflejo de verdaderos cambios
estructurales.
5.- Aunque la mayoría de las hipertrofias musculares probablemente son
el resultado de un incremento en el tamaño de fibras musculares individuales
(hipertrofia de fibras), algunas pruebas sugieren que puede intervenir también
un incremento en el número de fibras musculares (hiperplasia).
6.- Los músculos se atrofian, lo cual significa que reducen su tamaño y
su fuerza, cuando se vuelven inactivos, como consecuencia de lesiones o la
falta de utilización.
7.- La atrofia comienza muy rápidamente si se interrumpe el
entrenamiento, pero éste puede reducirse, como por ejemplo en un programa de mantenimiento,
sin que ello provoque la atrofia.
8.- Un tipo determinado de fibra puede adoptar características del tipo
de fibra contrario como reacción al entrenamiento, y hay pruebas que indican
que puede convertirse en este otro tipo de fibra como consecuencia de una
inervación cruzada o de una estimulación crónica.
---------------------
El tipo de contracción tiene importancia a la hora de las adaptaciones
al entrenamiento. Dado que se necesita menos ATP para el desarrollo de una
fuerza cuando las fuerzas externas alargan el músculo, en el ejercicio excéntrico el número de unidades
motoras activas disminuye y las necesidades energéticas son menores. Sin
embargo, debido al hecho de que la fuerza ejercida sobre las unidades motoras
activas es mayor en el ejercicio excéntrico, las contracciones de este tipo
producen un intervalo de respuestas retardadas después del ejercicio que
implican lesión muscular. Aparece debilidad y dolor (agujetas al primer día),
inflamación y edema (con un máximo de 1-3 días) y niveles plasmáticos elevados
de enzimas intramusculares (de 2-6 días). La reacción de fase aguda produce
activación del complemento, aumento de las citocinas circulantes, movilización
de los neutrófilos y redistribución de oligoelementos.
La ligera lesión muscular inducida por la contracción excéntrica y su
consiguiente respuesta puede ser el estímulo esencial para la hipertrofia
muscular.
Una variable importante a considerar es la velocidad máxima de
acortamiento, que viene determinada por la velocidad con que la miosina es capaz de
interaccionar repetidamente con los filamentos delgados. Es decir, de la
velocidad con que la miosina es capaz de unirse a la actina, producir el
desplazamiento del filamento delgado, desprenderse del ADP y del Pi, fijar nuevamente
ATP y separarse del filamento delgado para iniciar nuevamente este ciclo. La
velocidad máxima de acortamiento, corregida para el efecto de la longitud del
músculo, depende de la actividad ATPasa, o velocidad con que la miosina
hidroliza el ATP y transforma la energía liberada en energía mecánica. Sabemos
que la actividad miosina ATPasa viene determinada, principalmente, por la clase
de isoforma de cadena pesada de la miosina expresada en la fibra muscular,
siendo la jerarquía de velocidades IIx>IIa>I. En músculo humano se han
observado diferencias en velocidad máxima de acortamiento entre fibras de hasta
más de 20 veces. El tipo de isoforma de cadena ligera de la miosina que se
expresa en las fibras musculares también puede influir en la actividad miosina
ATPasa, pero en menor medida. El tipo de isoformas de la miosina presentes en
la fibra muscular viene determinada por la solicitación mecánica y, por lo
tanto, varía con el entrenamiento.
----------------------
La unión neuromuscular de la
fibra blanca tiene una terminación axonal más plana y elongada. Las vescículas
axioplásmicas son cerradas y los pliegues de union son mas largas y numerosos.
Estas diferencias ultraestructurales sugieren una mayor superficie potencial
para la interacción de las superficies axonales y sarcoplásmicas de las uniones
neuromusculares de las fibras musculares blancas comparadas con las rojas
•El músculo solo aumentará de masa cuando sus fibras adquieran mayor
espesor, lo que depende de la creación de miofibrillas adicionales.
•El estrés mecánico pone en marcha proteínas de señal que activan los
genes responsables de la síntesis muscular de mas proteínas contráctiles.
-Los núcleos del interior de las fibras no pueden dividirse.
-Los nuevos núcleos provienen de células satélite.
-Cuando se tracciona fuertemente el músculo, el área dañada atrae a las
células satélite que se incorporan al tejido muscular y comienzan a producir
proteínas para rellenar el espacio.
-Con tales núcleos supernumerarios, la fibra sintetizará más proteínas
y creará nuevas miofibrillas.
•Para fabricar proteínas, las células ejecutan el programa cifrado en
los genes, donde se indica el orden de engarce de los aminoácidos componentes.
-La información de
un gen (codificada en ADN) se copia en una molécula de ARN mensajero.
-Luego, el ARNm
lleva esta información a los ribosomas fuera del núcleo.
-Los ribosomas
ensamblan a.a. en proteínas (en actina o en una de las isoformas de miosina),
de acuerdo con las especificaciones del ARNm (traducción).
•Cuando se produce atrofia muscular, disminuye la cantidad relativa de
la isoforma lenta de miosina, incrementándose la cantidad de isoformas rápidas
de esta.
•Cuando los músculos sanos se someten a cargas pesadas de forma
repetida (entrenamiento de pesas), el número de fibras rápidas IIx desciende, y
se convierten en IIa. En estas fibras el núcleo deja de expresar el gen IIx y
comienza a expresar el IIa. Tras un mes de entrenamiento, todas las fibras IIx
pasarán a IIa. (Parece que el IIx constituiría una especie de normalización por
defecto)
•Cuando se abandona el ejercicio, las fibras IIa se convierten en IIx,
pero con un fenómeno de sobreexpresión, de modo que la cantidad de IIx será
superior a la inicial.
----------------------
Las características funcionales
del músculo dependen de las superficies relativas ocupadas por las fibras
lentas en la sección muscular. Cuanto mayor sea la superficie relativa ocupada
por las fibras rápidas, tanto más veloz será el músculo.
Tipo
I Tipo IIa Tipo IIx
Corredor maratón 4.800 nm2 4.500 nm2 4.600 nm2
Corredor velocidad 5.000 nm2 7.300 nm2 5.900 nm2
-----------------------
•Las transformaciones difíciles de obtener por entrenamiento (por
ejemplo, las de fibras IIa en fibras I), se lograrán muy pronto a través de
manipulaciones genéticas.
•Se estudia la expresión de los genes de miosina que, sin embargo, no
se expresan naturalmente. Son vestigios archivados de ciertos tipos de miosina
que debieron dotar a nuestros antepasados de un tejido muscular muy rápido para
escapar de los depredadores. (Tipo IIb.)
----------------------
El músculo, según las características de las fibras y estructuras que lo
componen presenta un comportamiento mecánico diferente al de otros materiales.
La viscoelasticidad determina que el comportamiento elástico quede comprometido
no sólo por la fuerza de deformación, sino también por el tiempo que tarda esta
en producirse. Es decir, se dice que un material es viscoelástico cuando la
deformación es dependiente de la carga aplicada y del tiempo en que se tarda en
aplicar.
Otro de los mecanismos que deben ser analizados es el nivel de
histéresis, que se manifiesta por el retraso del efecto sobre la causa que lo
produce. Representa la proporción de energía tensional que es disipada por la
amortiguación viscoelástica interna en cada ciclo de extensión, pudiéndose
calcular a partir del cociente entre el área que comprende la curva de tensión
(energía disipada) y el área que aparece por debajo de la curva de carga
(energía generada).
El reclutamiento diferente de las fibras de contracción lenta (ST) y
rápida (FT) influyen en el aprovechamiento de la energía elástica desarrollada
durante la carrera

----------------------
1.- La inflamación muscular aguda se produce hacia finales de una serie
de ejercicios y durante el período inmediato de recuperación.
2.- El inicio retrasado de la inflamación muscular se inicia un día o
dos después de la sesión de ejercicio. La acción excéntrica parece ser el
causante principal.
3.- Las causas propuestas de inicio retrasado de la inflamación muscular
puede ser el daño estructural en las células musculares y reacciones
inflamatorias dentro de los músculos.
4.- El modelo de secuencia de acontecimientos que produce la inflamación
muscular sería:
* daños estructurales
* deterioro de la
disponibilidad de calcio que conduce a la aparición de necrosis
* acumulación de
irritantes
* actividad macrófaga
aumentada
5.- La inflamación muscular puede prevenirse o minimizarse mediante:
* la reducción del
componente excéntrico de la acción muscular durante el inicio del entrenamiento
* el inicio del
entrenamiento con una baja intensidad incrementándolo progresivamente o
* comenzar con una
serie exhaustiva de elevada intensidad, que producirá mucha inflamación
inicialmente, pero que reducirá el dolor futuro.
